Amazing. I never heard half of this paper put out by Lamming’s lab. I copied some snippets below:
https://www.sciencedirect.com/science/article/pii/S1550413124003279
This review will focus on nutritional modulation of senescence, as exciting new evidence suggests aging and age-related diseases driven by senescence can be regulated by diet.
CR for 18 weeks decreases circulating senescence biomarkers in aged individuals. CR protects against senescence in multiple tissues of mice, rats, and humans.
Dietary protein and specific amino acids promote senescence. Methionine, arginine, BCAA. Taurine, proline reduce senescence. While there have been only a few studies examining how protein effects senescence, a recent study found that expression of p16 and p21 in the liver is associated with protein consumption,153 and PR reduces IL1α and TNFα in the livers of old male mice.154
Simple sugars, such as glucose, fructose, and galactose, promote senescence in culture and mice. Complex starches may be able to protect against senescence.
Increased dietary fat promotes senescence in multiple tissues of multiple model organisms to humans.
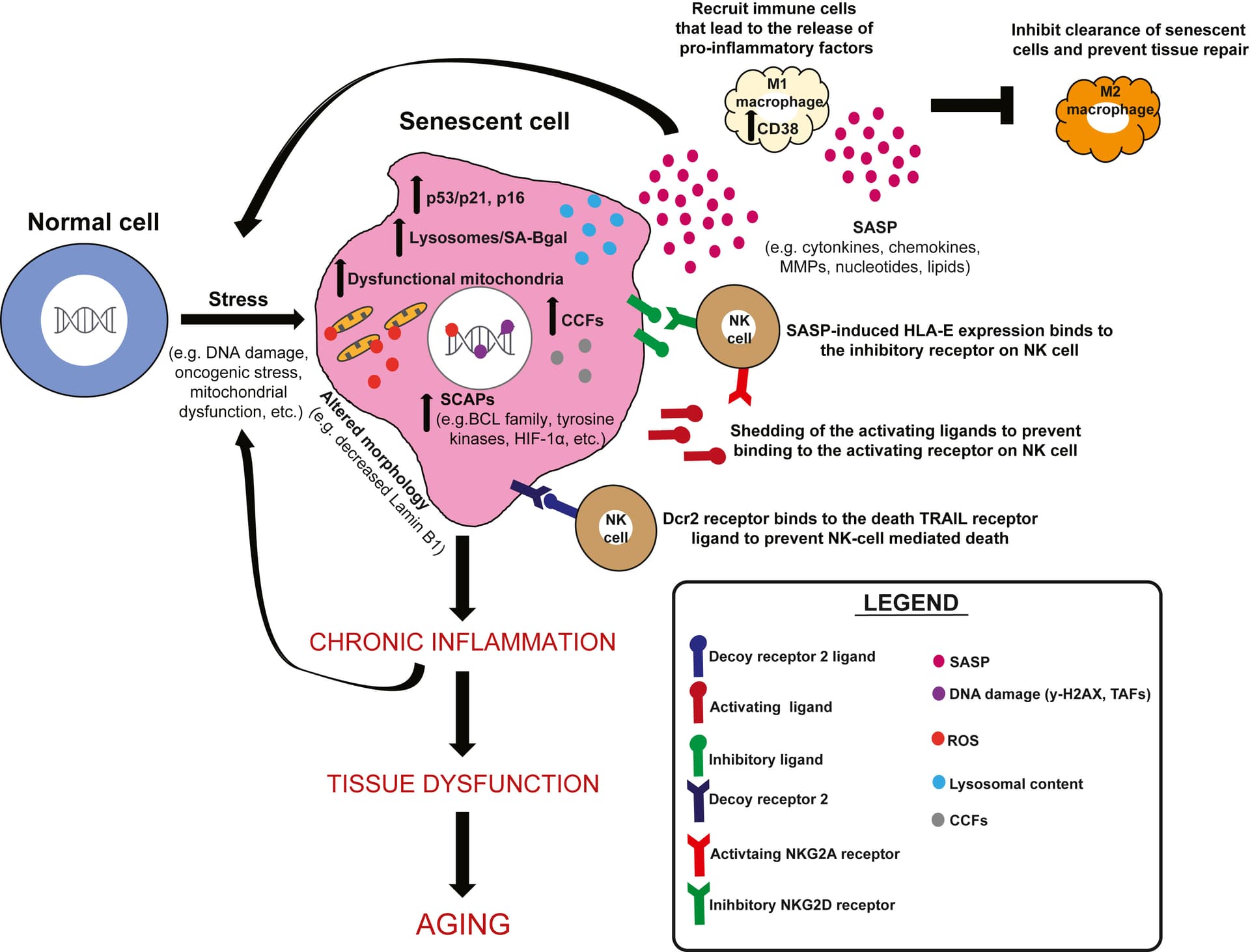
Micronutrients also play a role in organismal aging and senescence, particularly with suppression of SASP.245 Vitamin E has been shown to delay senescence in human umbilical vein endothelial cell (HUVEC) and fibroblast culture.246 Vitamin D decreases NF-κB activity in murine macrophage cells, suggesting it may reduce SASP.247
MR decreases senescence,156,157,169.
Outside of vitamins, polyphenols derived from cocoa have been shown to inhibit senescence via the modulation of sirtuins in auditory cell lines.249 Procyanidin C1, which is a polyphenolic component of grape seed extract (GSE), reduces SnC burden and increases the healthspan and lifespan of mice.250,251 In terms of minerals, chronic exposure to high-dose zinc can also accelerate senescence in human endothelial cells.252 Magnesium deficiency accelerates senescence in cultured cells,253
Protein and glucose restrictions have also been shown to activate AMPK.54,313 By contrast, high-fat diets inhibit AMPK.220,314Downregulation of AMPK activity induces premature senescence, while AMPK activation prevents premature senescence in MEFs, NIH3T3 cells, dental follicle cells, tendon stem/progenitor cell, human keratinocytes via maintenance of metabolic homeostasis, and murine skin.315,316,317,318,319,320 Conversely, AMPK induces senescence in endothelial cells, colorectal carcinoma cells, and fibroblasts.268,321,322 Persistent activation of AMPK accelerates senescence in a p53-dependent manner.323
IGF-1 itself also induces senescence in human fibroblasts, MEFs, 2BS cells, and mouse hepatic stellate cells in a p53-dependent manner327,328,329,330,331 but may decrease senescence in endothelial progenitor cells from older humans.332
The dietary level of specific macronutrients regulates mTORC1 activity, with restriction of protein or BCAAs leading to lower mTORC1 activity.148,152,343,344 Thus, nutrients can impact mTORC1-mediated senescence. Acquisition of senescence leads to constitutive upregulation of mTORC1 in human fibroblasts, making it resistant to both serum and amino acid starvation.345 High-protein diets, which promote mTORC1 activity in vivo in mice, increase levels of senescence in the liver of C57BL/6 male mice.153 In human HepG2 cells, BCAA supplementation enhances premature senescence via mTORC1.174 In addition, 16-month-old male C57BL/6J mice given dietary BCAA supplementation for 12 months had increased pro-inflammatory gene expression in visceral WAT,158 some of which overlap with common SASP markers. Consistent with this, BCAA accumulation within the cell promotes mTORC1-mediated SASP production in HEK293T cells.172
Other dietary macronutrients have been shown to activate mTORC1 activity, including carbohydrates. For example, in the context of pancreatic ductal adenocarcinoma, fructose induces mTORC1 activity.346 Galactose also induces mTORC1 activity in MSCs and rodent skin.196,197,198 Similarly, fats activate mTORC1. High-fat diets reduce AMPK phosphorylation, leading to increased mTOR activity
to maximize the therapeutic benefits of current senotherapeutics, diets should be taken into consideration. For example, rapamycin, which inhibits mTORC1 and extends lifespan, has been shown to have greater molecular effects in the context of high-protein diets, while metformin has the greatest molecular impact in the context of low-protein diets.344 This suggests that the drug efficiency may be altered by the diet consumed. While the efficacy of senolytics has been tested in high-fat diets, their interactions with dietary protein or carbohydrates have not been examined.48,207,213,396
it is important to consider the diets in which these individuals are consuming to maximize the benefits. For example, rapamycin, which acts as a senomorphic,402 might have the biggest benefits for those eating a high-protein diet, and negligible benefits for individuals eating a low-protein diet.