Thanks @Barnabas , great thread! Led me to investigate more as this looks like a critical process to understand. And it is directly related to @John_Hemming 's focus on the citrate cycle, ACLY and acetyl-CoA. From the study in the lead-off post:
Citrate metabolism controls the senescent microenvironment via the remodeling
of pro-inflammatory enhancers
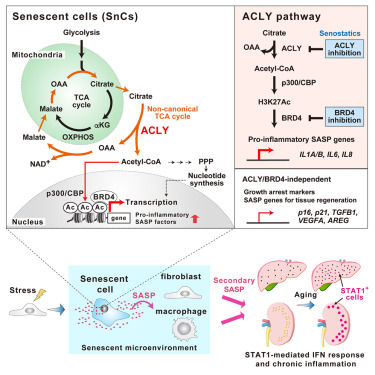
Here, we show that ATP-citrate lyase (ACLY), a key enzyme in acetyl-coenzyme A (CoA) synthesis, is essential for the pro-inflammatory SASP, independent of persistent growth arrest in senescent cells. Citrate-derived acetyl-CoA facilitates the action of SASP gene enhancers. ACLY-dependent de novo enhancers augment the recruitment of the chromatin reader BRD4, which causes SASP activation. Consistently, specific inhibitions of the ACLY-BRD4 axis suppress the STAT1-mediated interferon response, creating the pro-inflammatory microenvironment in senescent cells and tissues. Our results demonstrate that ACLY-dependent citrate metabolism represents a selective target for controlling SASP designed to promote healthy aging.
I started my search here:
https://en.wikipedia.org/wiki/ATP_citrate_synthase
Pharmacology
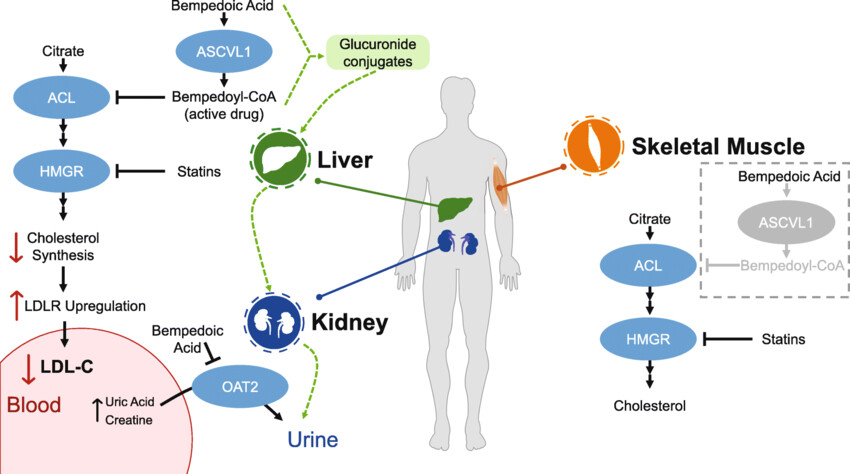
The **ATP citrate lyase (ACLY)**enzyme’s action can be inhibited by the coenzyme A-conjugate of bempedoic acid, a compound which lowers LDL cholesterol in humans.[14] The drug was approved by the Food and Drug Administration in February 2020 for use in the United States.
And the paper posted by @SilentWatcher is quite interesting and deserves further quotes - it’s a very long paper.
Abstract
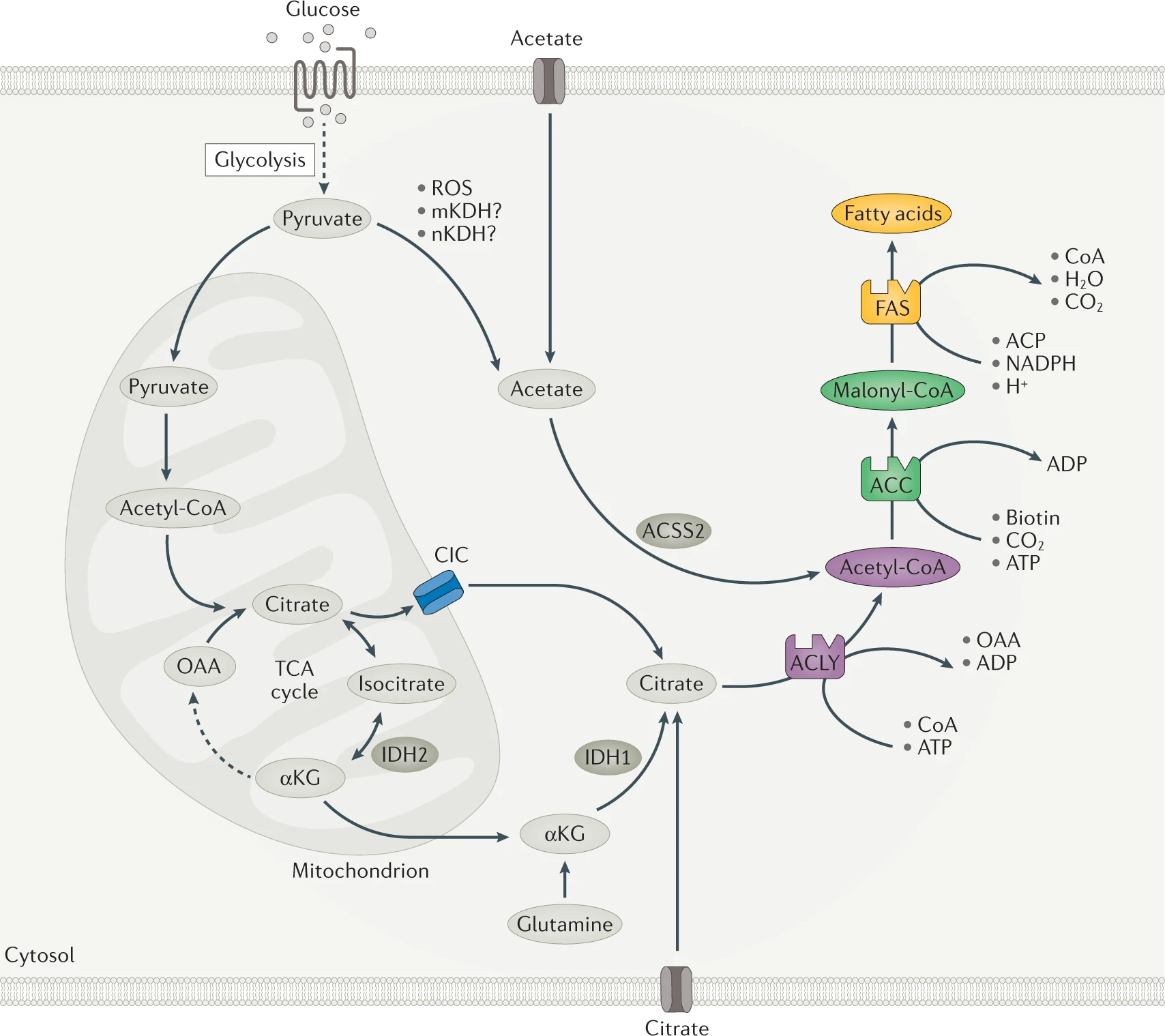
Fatty acids are essential for survival, acting as bioenergetic substrates, structural components and signalling molecules. Given their vital role, cells have evolved mechanisms to generate fatty acids from alternative carbon sources, through a process known as de novo lipogenesis (DNL). Despite the importance of DNL, aberrant upregulation is associated with a wide variety of pathologies. Inhibiting core enzymes of DNL, including citrate/isocitrate carrier (CIC), ATP-citrate lyase (ACLY), acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS), represents an attractive therapeutic strategy. Despite challenges related to efficacy, selectivity and safety, several new classes of synthetic DNL inhibitors have entered clinical-stage development and may become the foundation for a new class of therapeutics.
Introduction
Fatty acids are essential for cell survival as they serve as key structural components of cell membranes and important signalling molecules. Fatty acids are also the most calorically dense form of energy storage with the conversion of excess glucose into fatty acids protecting against glucotoxicity and providing a much larger energy reserve than glycogen for times of nutrient scarcity. Given the vital roles of fatty acids, cells have evolved mechanisms to maintain them at adequate levels. This includes mechanisms to take up exogenous fatty acids but also to generate fatty acids from alternative carbon sources through a series of enzymatic reactions, a process highly conserved across phyla known as de novo lipogenesis (DNL)1.
DNL is initiated when excess substrate availability, relative to cellular energy demands, leads to increases in mitochondrial citrate, which is exported from mitochondria into the cytosol by the mitochondrial citrate/isocitrate carrier (CIC; also known as CTP and SLC25A1) (Fig. 1). This cytosolic citrate is then converted into fatty acids by a series of biosynthetic reactions catalysed by ATP-citrate lyase (ACLY), acetyl-CoA carboxylase (ACC; also known as ACACA) and fatty acid synthase (FAS; also known as FASN). The expression of these enzymes differs across tissues and stages of development (for example, proliferation or quiescence). Expression and activity are also acutely and chronically regulated through transcriptional control and post-translational modifications that are linked to nutritional status (for example, fasting and feeding) and substrate availability (for example, fatty acids suppress DNL).
Although DNL is vital to maintain whole-body and cellular homeostasis, chronic elevations are associated with the development of a broad spectrum of diseases and disorders including cardiovascular disease (CVD)2,3, nonalcoholic fatty liver disease (NAFLD)4,5, type 2 diabetes (T2D)5,6, numerous cancers7,8, viral infections9,10, autoimmune diseases11,12, acne vulgaris13, neurodegeneration14 and ageing15. This suggests that pharmacological inhibition may be beneficial across multiple disease areas (Box 2; Supplementary Fig. 1). Several natural products have been identified as inhibitors of DNL and these have been adopted as a cornerstone for the development of synthetic inhibitors that display improved bioavailability, efficacy and specificity.
Bempedoic acid and BMS-303141, two of the better-characterized ACLY inhibitors, have strengthened a potential connection between weight loss and ACLY, with both reducing body weight gain and adiposity independently of changes in food intake in preclinical models160,166,167,172. Importantly, recent evidence has emerged from pooled analyses of clinical trials that bempedoic acid elicits modest weight loss in humans173. Studies examining the potential mechanisms by which ACLY inhibitors exert weight loss are warranted.
Bempedoic acid also reduced hepatic triglycerides and markers of inflammation in Ldlr−/− mice fed a diet high in fat and cholesterol. Importantly, the liver lipid-lowering effects of bempedoic acid are independent of liver AMPK activation167. In multiple mouse models, bempedoic acid also reduced fasting glucose, fasting insulin and glucose intolerance, suggesting improvements in insulin sensitivity172. Importantly, these effects appear to be translated to humans, as a meta-analysis of randomized trials suggests that bempedoic acid reduces new incidence or worsening of diabetes176. Whether bempedoic acid is effective at reversing NASH and fibrosis remains to be determined.
Bempedoic acid also suppresses hepatic cholesterol and fatty acid biosynthesis172 and its hypolipidaemic actions have been demonstrated in hyperlipidaemic hamsters172, obese Zucker rats166 and in mice deficient for ApoE167 or the LDL receptor179 in which atherosclerosis is also reduced. In humans, bempedoic acid promotes dose-dependent LDL-cholesterol lowering effects as monotherapy, and when combined with a statin or ezetimibe180. Unlike rodents, in which bempedoic acid has a profound effect in reducing both plasma triglycerides and cholesterol, the primary effect of bempedoic acid in humans appears to be a reduction in plasma LDL-cholesterol180. The lack of effect on plasma triglycerides may potentially be due to lower liver DNL in humans compared with rodents. In addition to lowering LDL-cholesterol, bempedoic acid also reduces several plasma markers associated with atherosclerotic CVD such as total cholesterol, non-HDL cholesterol, plasma apoB, LDL particle numbers and high-sensitivity C-reactive protein
In the past 50 years tremendous progress has been made in understanding the biochemical mechanisms and physiological significance of DNL in regulating cellular metabolism and whole-body energy homeostasis. Important steps along the way have included biochemical identification of the key metabolic intermediates in the conversion of glucose to fatty acids, the molecular cloning of the key enzymes regulating the process and the discovery of crucial allosteric and covalent mechanisms that regulate flux through the pathway. Subsequent studies in genetically modified mice revealed the physiological role of DNL, broadening our understanding of the complex connections that exist between multiple cellular processes and tissues well beyond the simple storage of excess calories in adipose tissue. Meanwhile, advances in structural biology laid the foundation for the molecular underpinnings by which natural products and new-generation small molecules inhibited enzyme activity. Although these compounds vary in their molecular target, chemical structure and physicochemical properties, the common action in which DNL is inhibited supports the notion that it is beneficial for the prevention and treatment of a broad spectrum of diseases.
Lastly, ACLY inhibition, which lowers acetyl-CoA, may affect the acetylation profile of many different histones, potentially having a wide array of differential effects on gene expression profiles and epigenetic programming. Thus, it is pivotal to consider the effects of metabolic intermediates when developing DNL inhibitors as a therapeutic approach. Similarly, genetic inhibition of ACLY in muscle and adipose tissue results in muscle weakness and lipodystrophy, respectively, but has been avoided by the development of the liver-targeted prodrug bempedoic acid. Liver-specific inhibition of DNL may also be helpful to avoid the potential detrimental effect of adipose tissue DNL inhibition on whole-body insulin sensitivity, given that adipose tissue DNL directly correlates with insulin sensitivity.
Ultimately, whether DNL inhibitors are safe and effective enough to be used as a monotherapy or will be used in combination with other therapies to enhance efficacy or offset liabilities, as has been proposed for ACC inhibitors in NASH, will depend on clinical trial results. In this regard, findings with the ACLY inhibitor bempedoic acid are encouraging as they suggest that chronic inhibition of DNL in the liver is safe when used alone or with other standards of care. Whether DNL inhibitors that target other cell types and organ systems become widely adopted as a cornerstone for treating other disease indications beyond CVD and NASH remains to be determined.